なぜヒトは老いて死ぬのか?

なぜ私たちは老いて死ぬのか?
多細胞生命にとっては長く生き、より多くの生殖期間を過ごしたほうが有利なはずです。
また社会性を持つ人間やゾウなどでは「長老」の存在が共同体の知識を蓄積する役割を果たすため、老いて死ぬシステムはその点、不利に働きます。
そのため長寿化が有利という理論は一見すると、生物学的にも社会学的にも妥当に思えてきます。
しかし長寿化の利点を享受する前提条件として、生物たちは種内の生殖競争に勝たねばなりません。
長寿の遺伝子を持っていたとしても、種内でモテなければ子孫を残せず、せっかくの長寿遺伝子も途絶えてしまいます。
より俗っぽい表現をするならば
「100歳まで生きるブサメンと50歳まで生きるイケメンがいた場合、子孫を残せるのは半分の寿命しかないイケメンのほう」
「60歳まで生きるイケメンと30歳までしか生きない超イケメンならば子孫を残せるのはさらに半分の寿命しかない超イケメンのほう」
となるでしょう。
(※ここでいうイケメンとは生殖能力の高さを比喩しただけのものです。生殖能力の高さとは、男女関係なく、妊娠のし易さや人生の早い時期から子孫を残す機会を獲得し易い特性を指します)
すると興味深い現象が起こります。
上の例はかなり乱暴ではありますが、長寿化が有利だとする理論が「生殖競争の結果」崩壊してしまうのです。
たとえ長寿であっても、その特性は繁殖真っ盛りの若い時期には影響しません。逆に歳を取ってから不利に働く特性(短命)は繁殖には影響しません。
米国のジョージ・ウィリアムズはかつて、この部分を真面目に考察し、生殖能力と寿命にかんする新理論「拮抗的多面発現仮説(きっこうてきためんはつげんかせつ)」を提唱しました。
なにやら難しそうな仮説ですが、その本質は「長寿のブサメンと短命のイケメン」の例えと同じです。
すなわち、長寿化という基本的に有利に働く個体の遺伝子が、生殖能力に全フリした個体の遺伝子に駆逐されてしまう可能性があるということです。
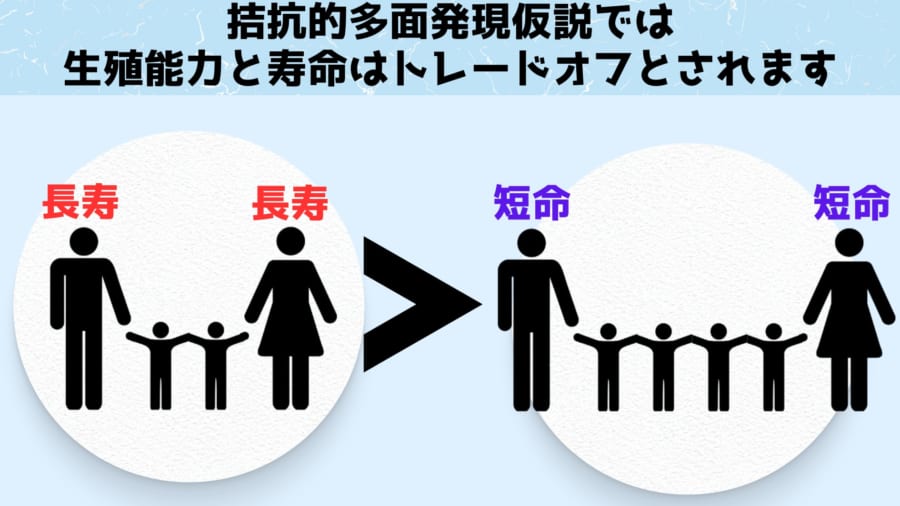
そのためウィリアムズは結果として「生殖能力と寿命のトレードオフ」が発生すると述べています。
現代的な遺伝学風に言うならば「生殖能力が有利になる代わりに、寿命には不利に働く遺伝子変異が起きても集団に受け入れられる」となるでしょう。
例えば、ある遺伝子は寿命に悪影響のある疾患に関連するが、生殖能力には有利に働くという特性を持っている場合があります。
このとき生物は、長寿において不利になるとしても、この遺伝子を優先的に継承してしまう可能性があります。
実際これまでの研究でも、ウィリアムズの拮抗的多面発現仮説はいくつかの生物種に当てはまることが示されています。
たとえばある昆虫の研究では、平均子孫数を「減らす」遺伝子変異が、虫の寿命を延ばしていることが示されました。
人間においても、冠状動脈疾患を起こす遺伝的変異を持つ人(短命の傾向)は、平均に比べてかなり多い子供の数を持つことが報告されています。
またフラミンガム心臓研究(FHS:心臓血管に関する大規模コホート研究)の報告によれば、女性の産んだ子供の数と女性の寿命の間に、有意な「負の相関」が観察されたことが示されています。
ただ、人間におけるこれらの報告がヒトという種全体において「生殖能力と寿命のトレードオフ」が存在する証拠となるかはまだ確かめられていません。
そこで今回、ミシガン大学の研究者たちはヒトにおいて拮抗的多面発現仮説が当てはまるかどうかを確認する、大規模調査を行うことにしました。