種の壁を一瞬で超えた衝撃映像

謎を解明するため研究チームは実験皿に用意したゼブラフィッシュの卵に、温度や培養液を調整したマウスの精子を加え、顕微鏡下でその挙動を観察しました。
すると予想どおり、マウス精子は魚の卵膜そのものには結合できませんでした。
しかし、しばらくすると精子の一部がゼブラフィッシュ卵の卵門がある極部に集まっていくのが確認されました。
精子は卵門の周辺にとどまるだけでなく、その小さな開口部に次々と進入し、卵膜の内部空間(卵膜下空間)にまで入り込んだのです。
マウス精子が魚の卵の門を物理的に通過しうることが示された瞬間でした。
この結果に、研究チームは大きな驚きを持って受け止めました。
第一著者のスマ・ガリボバ氏は「マウスの精子がゼブラフィッシュの卵門をくぐり抜ける瞬間を目の当たりにしたときは、信じられない思いでした」と振り返っています。
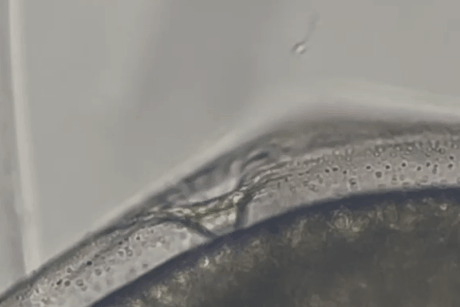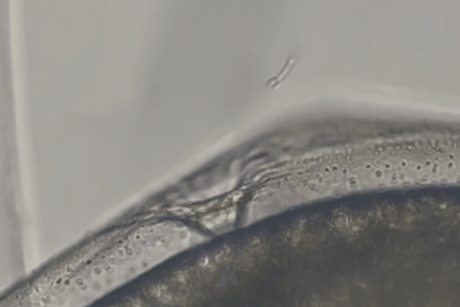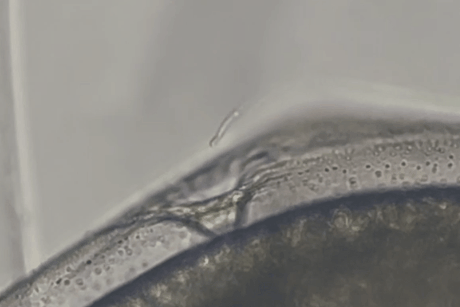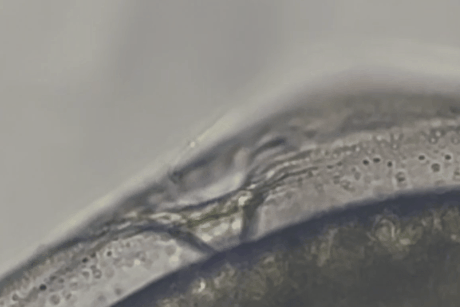
また実験では、マウス精子がゼブラフィッシュ卵に到達してから卵門に侵入するまで、およそ数十秒程度という短時間であることも示されました。
(※平均値は到達まで 48 ± 10 秒、侵入まで 24 ± 7 秒でした)
では、マウス精子はどのようにして魚の卵門を「探し当てた」のでしょうか。
その手がかりとして研究者たちは、卵門周辺に存在するとされる精子誘引物質に注目しました。
ゼブラフィッシュの卵膜表面を特定のレクチン(WGA-633)で染色する実験により、卵門の位置に糖タンパク質由来の物質が集まっていることが確認されました。
そこで卵膜のその物質を酵素(トリプシン)処理で除去すると、ゼブラフィッシュ由来の精子だけでなくマウス精子も卵門周囲にほとんど集まらなくなったのです。
つまり、マウス精子が魚の卵門を認識し近づくには、卵門部位に存在する何らかの誘引因子(研究では仮に「MP」と呼称)が必要であることが示唆されました。
さらに詳しく解析するため、研究チームはこの誘引因子が作り出す濃度勾配と精子の位置との関係を調べました。
その結果、卵膜上で卵門に向かうほど当該物質の濃度が高くなり、実際にマウス精子の数も卵門に近づくほど増加していることが確認されました。
これは、卵門の誘引因子が精子を卵門へ導く化学的な道しるべ(キモアトラクト)として機能している可能性を強く示すものです。
また、マウス精子が卵門を通過した後の挙動についても重要な発見がありました。
魚類の精子には存在しない先体(アクロソーム)という酵素の詰まった「帽子」がマウス精子の頭部にはありますが、この先体が放出される先体反応は本来、哺乳類の精子が卵子と出会ったときに起こる現象です。
研究チームが先体部分に赤い蛍光標識(mCherry)を持つトランスジェニック・マウスの精子を使って観察したところ、ゼブラフィッシュ卵門を通過して卵膜内に入ったマウス精子の多くは先体が未反応のまま残っていることがわかりました。
つまり、魚の卵の環境ではマウス精子の先体反応が十分に起こらず、精子は酵素を放出しないまま内部に存在していたのです。
この理由は定かではありませんが、ゼブラフィッシュの卵は元来アクロソームを持たない精子に対応した仕組みを進化させてきたため、マウス精子に先体反応を促すシグナルを与えられない可能性があります。
事実、今回マウス精子は卵膜内空間に進入したものの、卵そのもの(卵細胞)との融合・受精には至りませんでした。
この点は「種の壁」の厳しさを示すものですが、逆に言えば卵門を通過するまでのプロセスについては種を超えて共通する部分があることが示されたといえます。
さらに注目すべきことに、マウス精子が魚の卵門を通過するにはCatSper(キャットスパー)と呼ばれる精子固有のカルシウムイオン・チャネルが不可欠であることも判明しました。
CatSperは精子のべん毛の活動性(ハイパーアクティベーションと呼ばれる力強い運動)を制御するタンパク質で、受精に重要な役割を果たします。
研究チームがCatSper1遺伝子を欠損した変異マウス(CatSper1 Nullマウス)の精子を用いて同じ実験を行ったところ、これらの精子はゼブラフィッシュの卵門にほとんど近寄らず、通過することもできませんでした。
つまり、精子が魚の卵門を見つけ侵入するためにはCatSperチャネルによる鞭毛運動の活性化が必要であり、その機能なしには種を超えた相互作用も起こらないことが示されたのです。




























![シルバーバック かわいい海の生きもの CUBE 2X2 キューブ ツーバイツー|海の生き物デザイン 立体パズル スピードキューブ 5cm 子ども〜大人向け 知育 ギフトに最適 ([バラエティ])](https://m.media-amazon.com/images/I/41EJOOLgGXL._SL500_.jpg)