



致死システムを維持する「進化のシナリオ」
なぜ第1染色体を大小の2種類に分け、両方がなければ生き残れないようにしたのか?
研究者たちは鍵となるのは先祖型の遺伝子を持つ近縁種であると述べています。
大小の染色体からなる致死システムを持っている集団に先祖型の集団が接触した場合、その子孫は全て混合型となり、第1染色体と大きな染色体(A)と小さな染色体(B)を全て持つことになります。
しかしこの場合、本来ならば第1染色体だけ、あるいは大きな染色体(A)と小さな染色体(B)1本ずつでよかったところに、余計な染色体が加わることになります。
染色体の数が多くなると、生命の設計図となる遺伝子の数も増え、結果として生産されるタンパク質の量に影響を与えて生存に不利になることがあります。
研究者たちはこの3種類の染色体を持つ子孫が受ける「ペナルティー」の大きさを操作し、どうなるかを追跡しました。
ここで言うペナルティーとは、生存率や雌の繁殖力、雄の魅力などが含まれています。
すると3本の多すぎる染色体を持つペナルティーが50%を超えると、3本の染色体を持つ個体が徐々に排除されていき、致死システムを持つ子孫のほうが優勢になっていきました。
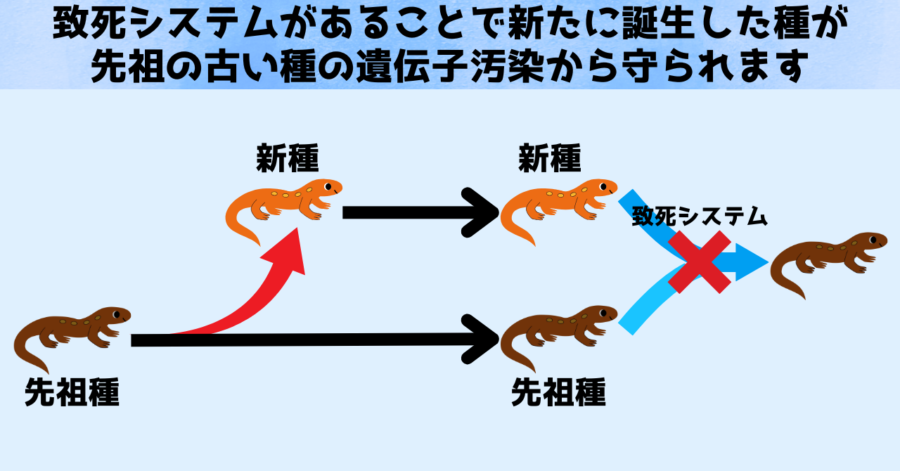
この結果は、致死システムを採用することで、先祖型の侵入によって自分たちの遺伝子が変化してしまうのを、保護できることを示しています。
致死システムがあることで、先祖型と異なる種として並行して生存しやすくなるのです。
また致死システムの出現は第1染色体内部の遺伝子の奪い合いというイベントが起きてから、2世代以内に確立されます。
さらにイモリの遺伝子を分析したところ、染色体内部に遺伝子の奪い合いを誘発させやすい構造(反復配列)が多数存在することが判明しました。
つまり、致死システムそのものの出現が生物にとって決して難しいことではなく、イモリにはもともと致死システムの元となる遺伝子の奪い合いを起こしやすい配列特性があり、致死システムを持つ種が出現した以降は先祖型との交雑を排除しやすくなるわけです。
ある意味で、致死システムとは先祖型と簡単に種分化ができるお手軽進化システムでもあるわけです。
ただこれの特徴はどれも致死システムの出現において短期的なメリットでしかありません。
このような短期的メリットが長期に渡る進化スケールでどれほど通用するのでしょうか?
謎を解明するため研究者たちはさらなるシミュレーションを行いました。
すると先祖型の染色体を持つ集団のサイズが500を超える条件では致死システムを持つイモリが1匹出現することが判明。
さらにシミュレーションを続けて先祖型から致死システムを持つ新種が誕生する確率を調べるため反復試行を行いました。
すると持続的な致死システムが形成される例がゼロではなく2%ほど存在することが確かめられました。
この結果は、先祖型の集団と差別化しようとする致死システムの出現は決して理不尽であり得ないことではなく、条件さえ適切ならば十分に出現可能であることを示しています。
もちろん、イモリたちの生息環境が激変して生存が困難になった場合、致死システムを持つ種は存続がより困難になるでしょう。
しかし増殖力だけが重要ならば、現在も地球は単細胞生物しか存在しない世界のままのはずです。
増殖力に多大な不都合がある種も、長年に渡る繁栄を享受し多様化も可能なのは、地球の環境が多様である証拠とも言えるでしょう。


























![シルバーバック 【釣って遊ぼう!サメまみれ!】 知育玩具 幼稚園 小学校 入園 入学 お祝い プレゼント 準備 ([バラエティ])](https://m.media-amazon.com/images/I/41ejNUfrJZL._SL500_.jpg)



























後半のシミュレーションが、とても面白いです。このようなdry lab は有益ですね。
前半がよくわかりません。最初のAとBあたりの話ですが、
①先祖の遺伝子型は、AA?BB?それともそれ以外?純系なのは分かるけど…。
②さらに、「3つ目が加わる」というのもわかりません。ダウン症のような状況?通常の「ヘテロ」の交雑なら、基本的に2本ですよね?たとえば、分化した、例えば、ABに先祖の型のAA(正しい仮定か分かりませんが)が接触すると、戻し交配的に、AAかABができるだけで、ABAなどにはならないのでは?
本文から読み取った範囲での回答ですが、前半のABは「大きな遺伝子A」と「小さな遺伝子B」で、これはかつては別な由来の遺伝子だったと考えられていたが、今は中くらいの遺伝子(祖先型。全ての情報を持つ)2本が遺伝情報を奪い合い、AとBになったと考えられています。なので祖先の遺伝子は(A+B)÷2の遺伝子です
後半の説明の為に(A+B)÷2=Cとします
祖先遺伝子CCと、進化したABが交雑すると、CAもしくはCBになります。CAもCBもC由来の過不足のない情報があるため成長は出来ますが、AかB由来の過剰な情報を持っているので、単純なC個体より遺伝情報が過剰になる部位が生じます。それが「3つめが加わる」という意味だと思います
前半部
おそらく、A=長くなった染色体、B=短くなった染色体って定義してるだけだから、先祖型はそうなる前の、同じ長さの相同染色体ってことじゃないかな。
後半部
同意。もしかしたら、
A=長くなった染色体
B=短くなった染色体
に加えて、
C=先祖型染色体
とすると、
AC、あるいはBCの染色体の組み合わせでは、必要なゲノムが足りずに発生がうまくいかないって話だろうから、染色体ABをもつ集団内に、3つ目の染色体Cが流入すると…ってことを言いたいのかな?
先祖がAAだとすると新種はBCってことじゃないの?
でも先祖との交配で1番染色体だけ3倍体ABCとはならないような気もする…
普通の交配で考えたらABとACができると思うけど…
AB,ACのどっちにしろ
均等配分のAがあるから発生自体はするけどB,Cの部分で発現量の均衡が崩れて
確実に障害を負うみたいな?
1番染色体ってことは一番大きな染色体ってことでこれがトリソミーなんて起こしたらまず発生すらできないと思う
合っているか分からないのですが、ABはお互いの遺伝子を奪い合っていてどちらも少しずつ変化しているので、正確には先祖型の染色体をAAとすると新種のヘテロはA’Bと表せ、AとA’は異なるため、どちらかが選択されるということはなくトリソミーになりやすいのではないでしょうか?
とても面白いですが、適応の説明が出来ていないような。
グループセレクションなんでしょうか?
染色体3本ではなく染色体2本のいずれかが3種類め(AでもBでもない先祖型)になると不利ということではないでしょうか。
記事中の例でいうなら孵化できる新種は「脳脳」と「脊髄脊髄」の染色体がペアになって脳×2脊髄×2のバランスのはず。
ここに先祖型の「脳脊髄」が組み合わさると脳×3脊髄×1か脳×1脊髄×3となってバランスが狂い、通常の新種に比べて生存や繁殖の成功率が下がることはありそうです。
解りやすい説明ですが図では先祖型が続いていくように見えます。新種が続いていくのではないですか。勘違いだったら済みません。
両方とも続いていくと思います。
このシミュレーションでわかるのは、致死システムによって先祖と新種の間で雑種が生まれることが妨げられるということだけです。
先祖が死滅するのか、新種が適応するのかは、この研究ではわからないと思います。
恐らくですが、先祖型をAとすると、収奪により長くなった方のAがAl(A long、本文内ではA)、奪取された方がAがAs(A short、本文内ではB)という言うことなのではないかと読みました。
このA Al Asの3つが異なる3つにあたるのではないかと。
元々AAだったが、変異によってAlとAsの遺伝子長の違う染色体ができた。AlとAsがホモになると、本文中の例えに従うなら脳3脊髄1の設計図となるため致死となる。ヘテロなら脳2脊髄2なので結果的には問題ない。
先祖型と交雑するとA Alか、A Asのいずれかの組み合わせにならざるを得ないため、設計図の過不足が起きるから致死となるが、先祖型との交雑を避ける機能を結果的に果たしてしまい分化を促進した。
元論文中では発現する部位(本文中では脳、脊髄)に対してA,Bと呼称し、染色体自体をA,Bとは呼称してないから、染色体に着目する場合はこれで良いと思うんですが…
一応元の論文も確認したから合ってると思うけど、自信ニキ助けて。
>お互いの遺伝子の奪い合いを行っていたことも判明します
生き物は卵の時点で生存競争しているのでしょうか
業が深いですね
イモリのたまごの死の要因について興味深く読ませていただきました。先祖型の500分の一の割合で致死システムが発現するとはおもしろいです。それらが混在しているときになんらかの環境的な要因で先祖型が死滅した結果、致死システムが生き残ったと考えるのが自然でしょうか。致死システムは他の遺伝子に汚染されにくいこと利点としている様ですが、それが死滅しなかった要因とどう関わっていたのか、その辺は今後の研究で明らかになったらいいですね(^^)
おそらくこういうことではないかと思ったのですが、
①ここでは、A:大きい遺伝子、B:小さい遺伝子で解説されているので、先祖型はどちらでもない変異前のそれぞれ同じ長さの遺伝子です。前のページに解説があります。
②仮に先祖型の遺伝子について、染色体ABとabの一対を持つとすると、
致死的システムの遺伝子は、変異のため染色体AaとBbの一対を持つことになります。
その場合、先祖型と致死的システムの交配の結果ではABとAa、ABとBb、abとAa、abとBbの組み合わせになります。本来なら2:2で一対×2本の染色体となるA(あるいはa)対B(あるいはb)の遺伝子割合が1:3の不均衡になり、トリソミーの状態になると思われます。