半分死ぬ仕組みは誤解されていた
多くのイモリでは親が産んだ半分の卵が自動的に死んでしまうという奇妙な致死システムが存在します。
そのためイモリは過酷な自然環境や捕食者から生き延びるだけではなく、種の存続のために必要とされる卵の2倍を産み続けなければなりません。
100個の卵のうち大人になって子供を残せるのが1個という条件ならば、単純計算をすると、親は死亡する分を補うため200個の卵を産まなければならなくなります。
生物学の常識では、このような致死システムはデメリットでしかなく、明らかに自然選択の理に反しています。
なぜイモリはわざわざ、卵が半分死ぬ仕組みを進化させたのか?
生物学者は長年にわたりイモリの奇妙な致死システムの謎を追い続けていました。
ですが逆に、謎はより深まります。
死んだ卵は親が食べて栄養補給するのだろうか?
それとも生まれてきた子供たちの食料になるのではないか?
いや、死ぬ卵は捕食者に対するダミーで中身は空なのではないだろうか?
実験室の環境が悪いだけで、自然環境ではちゃんと全部孵化するのではないか?
長年の研究により、このような簡単な説明はかなり早期に排除されてしまったからです。
この結果は、卵が半分死ぬ仕組みはイモリの生殖出力にとって、全く補償されない、純粋な損失でしかないことを示しています。
意味のない死というわけです。
しかし死ぬ意味はわからなくても、死ぬ仕組みの解明は続いていました。
1980年代に行われた研究では、この仕組みが第1染色体の異常であることが判明します。
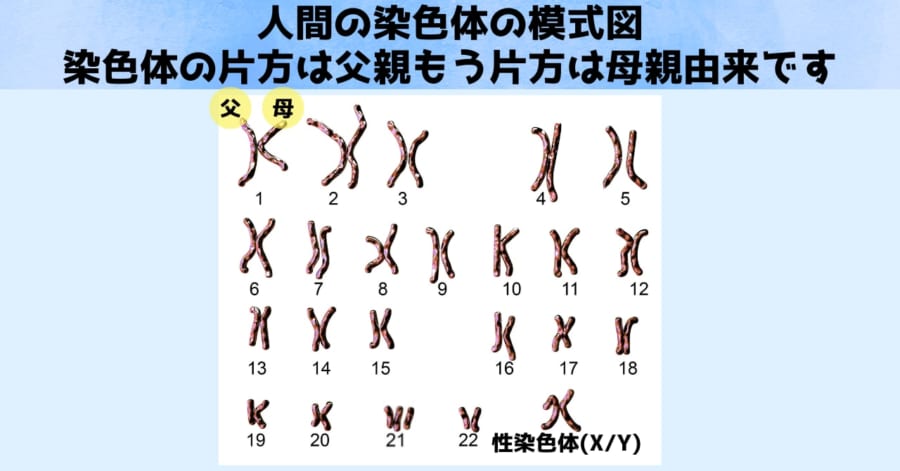
人間を含む多くの動物は、父親と母親から染色体セットを1つずつ受け継ぐことで、2つのコピーを持っています。
通常、父親由来と母親由来の染色体は同じ大きさであり、違うのは性染色体だけです。
しかしイモリの場合、第1染色体には大きいものと小さいものが存在していることがわかりました。
そして大きい染色体(A)を2つ、あるいは小さい染色体(B)を2つ受け継いだものは死んでしまいます。
生き残るのは、大きい染色体(A)と小さな染色体(B)を1本ずつ受け継いだ卵だけになります。
そのため以前には、小さい染色体(B)しかない場合は遺伝子不足によって死亡し、大きい染色体(A)しかない場合は遺伝子過多によって死亡すると考えられていました。
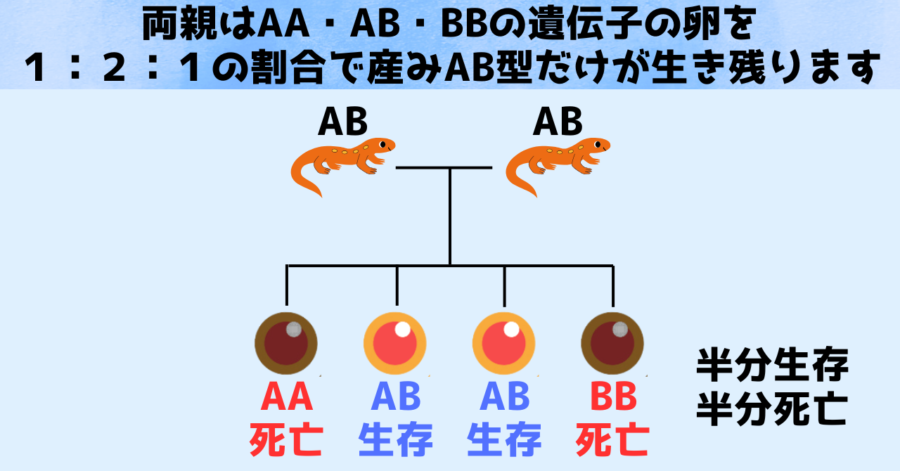
上の図では大きい染色体(A)を2本持つAA型と小さい染色体(B)を2本持つ卵が死に、大きい染色体(A)と小さい染色体(B)を1本ずつ持つ卵だけが生き残る様子を描いています。
中学生で習うメンデルの遺伝法則を第1染色体のみで再現した状態と言えるでしょう。
(※ショウジョウバエなどの研究を行っている研究室では、この致死システムを人工的に再現することで、有害な突然変異の系統を安定的に維持するために使用されています)
また驚くべきことに、この仕組みは均衡致死システムとして知られ、イモリ属の1つTriturus属の9種類全てにみられる特徴となっています。
さらに同じような致死システムは、イモリの異なる種で独自に獲得された可能性も示されています。
本当に不利なシステムならば、種の分岐が起きる前に排除されたり、たまたま採用された1回だけのレアケースになってもよさそうなものですが、そうではなかったのです。
これまでの有力な説では、大きな染色体(A)と小さな染色体(B)はもともとは系統の異なる性染色体であり、致死的なシステムは段階を踏むようにして徐々に形成されていったとされていました。
卵が半分死ぬような理不尽なシステムが急に出現しても、あっという間に絶滅してしまうため、変化は少しずつ進行していったとする説です。
また系統の異なる性染色体であれば、そもそもの大きさが異なっていても説明がつきます。
ただこの説を証明するには、染色体に対する詳細な分析が必要です。
そこで今回ライデン大学の研究者たちは、致死システムを採用するイモリの染色体に対して4226個ものDNAマーカーを打ち込み、詳細な遺伝子地図を構築しました。
また同時に致死システムを採用していないイモリの染色体に対しても同様の遺伝子地図を構築し先祖の代理として扱います。
そして両者の遺伝子地図の比較を行いました。
すると致死システムの獲得は徐々に行われたのではなく、1度の突然変異がキッカケになっていることが判明します。
この結果は、既存の有力な説に反するものです。
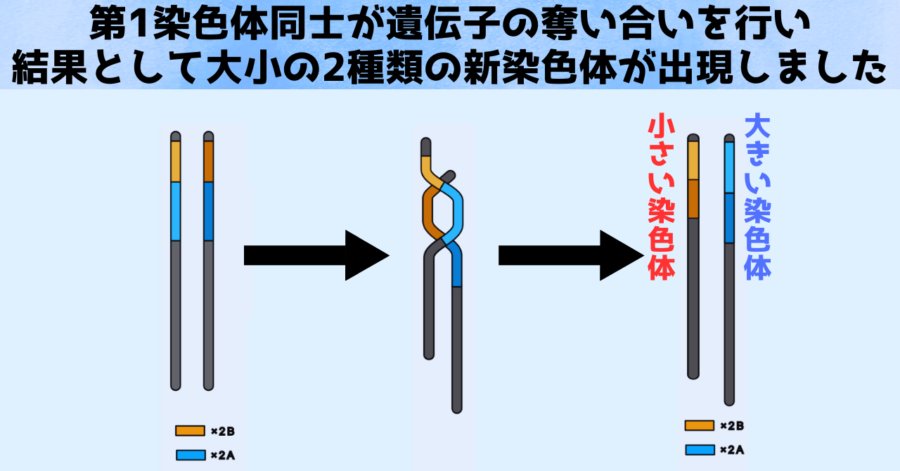
さらに第1染色体に対する分析を行ったところ、大きな染色体(A)と小さな染色体(B)はお互いの遺伝子の奪い合いを行っていたことも判明します。
上の図はその過程を示したものになります。
大きな染色体(A)と小さな染色体(B)はもともと同じ先祖型でしたが、遺伝子を奪い合った結果、大きな染色体(A)はA連鎖遺伝子(寒色)と呼ばれる領域を獲得し、小さな染色体(B)はB連鎖遺伝子(暖色)と呼ばれる領域を獲得しました。
一方の染色体が他方より大きいのは、この奪い合いが同等の割合ではなく不均衡であったことが原因だったのです。
さらにこれらの連鎖遺伝子たちは、相手に存在した自分と同じ遺伝子を多数含んでいることが判明しました。
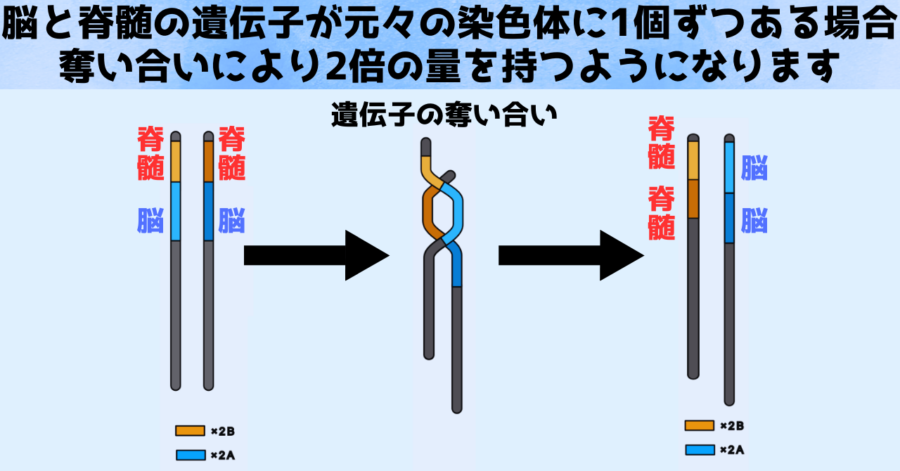
たとえば父親由来の染色体と母親由来の染色体の両方に、脳にとって重要な大きな遺伝子と脊髄にとって重要な小さな遺伝子が1つずつ含まれていたとします。
ですが染色体同士の間で奪い合いが起こった結果、大きな染色体(A)は脊髄の遺伝子を失う代わりに脳の遺伝子を2重に獲得し、小さな遺伝子(B)は脳の遺伝子を失う代わりに脊髄の遺伝子を2重に獲得したとします。
すると、脳の遺伝子のほうが大きかったため、染色体の大きさに違いもうまれます。
(※脳と脊髄はたとえであり、実際にはより多くの遺伝子の奪い合いが起きています)
またイモリは脳を備えた脊椎動物ですので、卵が正常に発達し生き残るには、脳の遺伝子も脊髄の遺伝子も両方必要になります。
そのため大きな染色体(A)だけしか持たない卵や、小さな染色体(B)だけしか持たない卵はどちらも遺伝子不足を起こして死んでしまったのです。
既存の説では小さな染色体(B)だけが遺伝子不足になると考えられていましたが、実際には大(A)小(B)どちらの染色体も遺伝子不足だったわけです。
研究では実際に、大きな染色体(A)と小さな染色体(B)のそれぞれにおいて、特定の遺伝子が2倍量存在していることが示されています。
逆に大きい染色体(A)が2本ある場合はなどは、特定の遺伝子が4倍体に近いというデータも得られています。
問題は、なぜこのような仕組みが続けられたかです。
同じ第1染色体が大きな染色体(A)と小さな染色体(B)の2種類に別れ、両方がなければ死んでしまう仕組みが解明されたとしても、卵の無駄を作るメリットまでは説明できません。
そこで研究者たちは、分析結果をもとに新たな進化のシナリオを描きました。