オタマボヤは「受精から生まれるまで3~4時間」「大人になるまで10時間」しかかからない:高速発生の仕組み
なぜオタマボヤはそんなに生き急いでいるのか?
謎を解明すべく研究者たちはオタマボヤの全発生過程にわたる遺伝子発現プロファイルを網羅的に解明し、近縁のホヤとの比較を行うことにしました。
具体的にはオタマボヤの受精卵から成体になるまでの各段階でどんな遺伝子が働いているかを網羅的に調べ、そのデータベースを構築するという壮大な計画です。
これにより、オタマボヤの発生プログラムの全貌と近縁種との異同が明らかになると期待されました。
すると、幾つか奇妙な結果が得られました。
1つ目は発生初期からの活発な遺伝子活性です。
受精後ごく初期(16~32細胞期)の胚で、既に約950個もの遺伝子が初期発現を開始していることが判明しました。
これは母親由来の物質だけに頼らず、胚自身のゲノムが非常に早い段階から動き出すことを意味します。
典型的な動物発生では受精卵由来のmRNAやタンパク質により初期の発生が進み、ある段階でゲノム活性化が起こります。
しかしオタマボヤではそのスイッチオンが極めて早期に起こるようです。
2つ目は転写因子群の大幅な簡略化でした。
転写因子は、細胞という巨大な図書館で“本棚の鍵”を持つ司書のような存在です。
読みたい遺伝子(本)の前に立ち、鍵を差し込んで表紙を開くと、その設計図のコピー機に送られて設計図の部分写し(RNA)が作られ始めます。
どの棚を開けるか、何冊同時に並べるかを瞬時に判断しているため、司書の采配ひとつで細胞の“今日の業務”が決まります。
もし鍵を失ったり間違った棚を開けたりすると、必要な設計図が読めずに細胞の機能全体が滞る――まさに細胞経営の要(かなめ)です。
そんな大切な転写因子ですが、32細胞期の胚で発現している転写因子は25種類に過ぎないことが分かりました。
同じ段階のホヤ胚ではこの倍以上の転写因子が働くことが知られています。
つまりホヤの胚発生を支える転写因子の半数以上が、オタマボヤでは存在しないか全く使われていないのです。
言い換えれば、オタマボヤはホヤに比べて発生過程の遺伝子制御網が大幅に圧縮・簡略化されています。
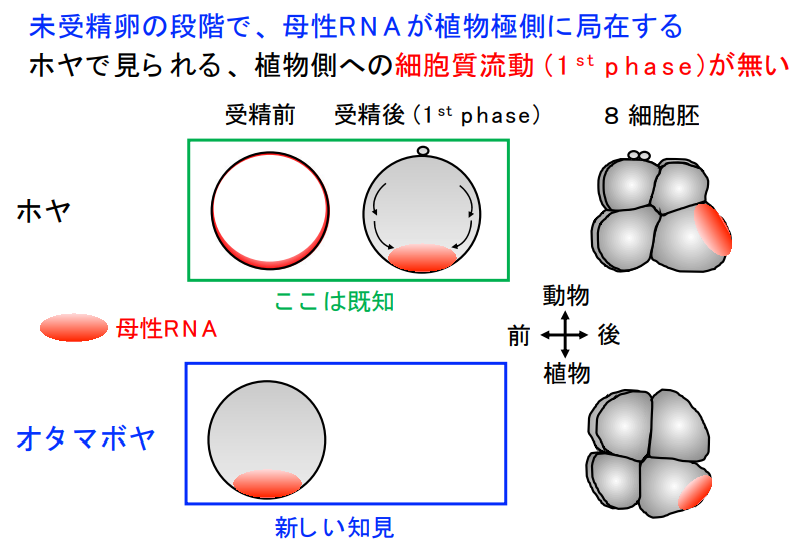
3つ目は母性mRNA局在パターンの違い
多くの動物では、未受精卵中に蓄えられた母性因子mRNAが受精後に胚内の特定領域へ再分配されることで体の前後が決められます。
卵子の中にはあらかじめ大量の設計図の部分写しが存在しており、受精が始まると「部分写し作業(RNAの0からの生産)」の過程をかなり飛ばして、素早く発生が始まります。
ホヤの卵は、受精するとすぐに中身が「ザバーッ」と流れ動き、母親が入れておいた遺伝子メッセージ(母性mRNA)が卵の下側に集まります。
ところがオタマボヤは、最初からそのメッセージを下側に並べたまま受精を待つので、あとでかき混ぜる作業がいりません。
さらに別のメッセージは、卵が分裂していく途中で“仕分け係”のように後ろ側の細胞へ振り分けられるしくみが働きます。
つまりオタマボヤでは、受精前から必要な母性因子の配置が完了しており、受精後の大掛かりな再配置を行いません。
その分、オタマボヤ胚は発生開始と同時に自前の遺伝子群を起動し、少ない調節因子で各細胞の運命を一気に決定して突き進みます。
まさに「一直線に突っ走る」タイプの発生プログラムであると言えるでしょう。
海の表層は栄養や温度が日替わりで変わる気まぐれな世界です。
オタマボヤは卵からわずか数日で大人になり産卵できる超時短ライフを武器に、「いい環境が整ったら逃さずに一気に増えることができる」戦略をとります。
プランクトンの大発生など短期間しか続かない餌の山を取りこぼさないので、わずかな好機でも個体数を爆発的に増やせます。
逆に環境が悪化すると成体はあっさり死に、水中に残った卵や幼生が漂いながら次のチャンスを待つため、大集団としてのリスクも抑えられます。
こうした瞬発力重視のライフスタイルこそが、変動の激しい外洋でオタマボヤが生き残る鍵になっています。




























![シルバーバック 【釣って遊ぼう!サメまみれ!】 知育玩具 幼稚園 小学校 入園 入学 お祝い プレゼント 準備 ([バラエティ])](https://m.media-amazon.com/images/I/41ejNUfrJZL._SL500_.jpg)