そのウイルスはエネルギーを作る遺伝子を持っていた
PelV-1とはいったい何者なのか? そして、あの「とんでもなく長いしっぽ」にはどんな意味があるのでしょうか?
その正体を探るために、研究チームはまず、このウイルスの見た目をじっくり観察することから始めました。
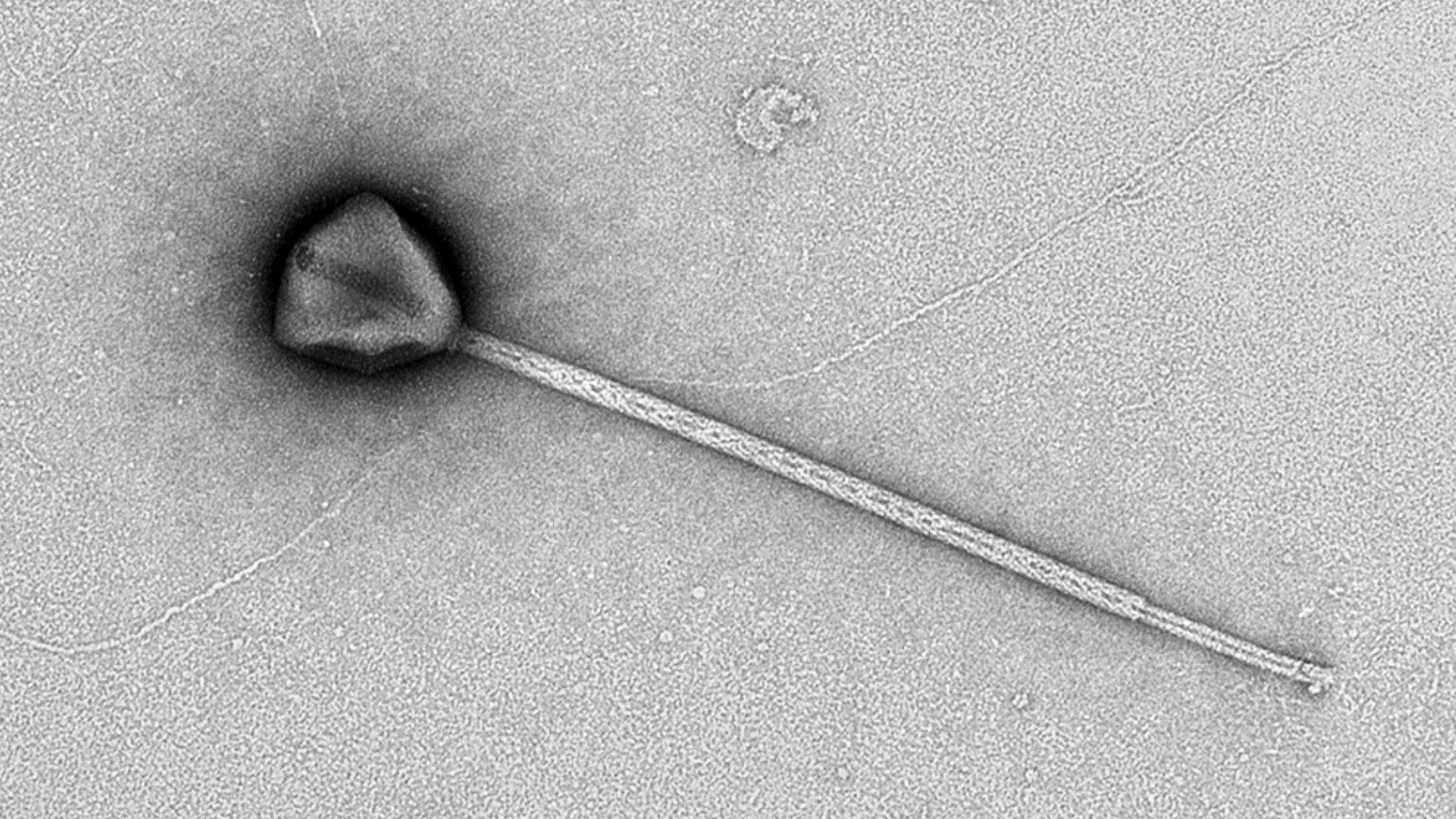
特殊な電子顕微鏡を使ってウイルスを見てみると、PelV-1は直径およそ200ナノメートル(0.2マイクロメートル)の多角形のかたい殻(カプシド)を持ち、そこから細長いしっぽのような構造が1本、ひょろっと伸びていることが分かりました。
このしっぽは幅がたった30ナノメートルくらいしかありませんが、長さはなんと最大で2.3マイクロメートルにもなるのです。
これは、カプシド本体の10倍以上の長さです。
大腸菌などの普通の細菌と同じくらいの長さだと考えると、いかに大きいかが分かります。
このしっぽは、今まで見つかったどのウイルスよりも長いとされています。
たとえば、P74-26というウイルスのしっぽは約0.875マイクロメートル、ツパンウイルスという巨大ウイルスでも0.55~1.85マイクロメートル。それらを上回る長さなのです。
さらに驚くのは、PelV-1のカプシドの反対側には、星の形をした「ふた」のような部分(スターゲート)と、その近くに短く太い突起があることです。
この突起がどんな役割を果たしているかはまだはっきりしていませんが、ウイルスの中身を放出する通路と関係しているかもしれないと考えられています。
続いて、研究チームはPelV-1の「中身」、つまり遺伝子の情報も解析しました。
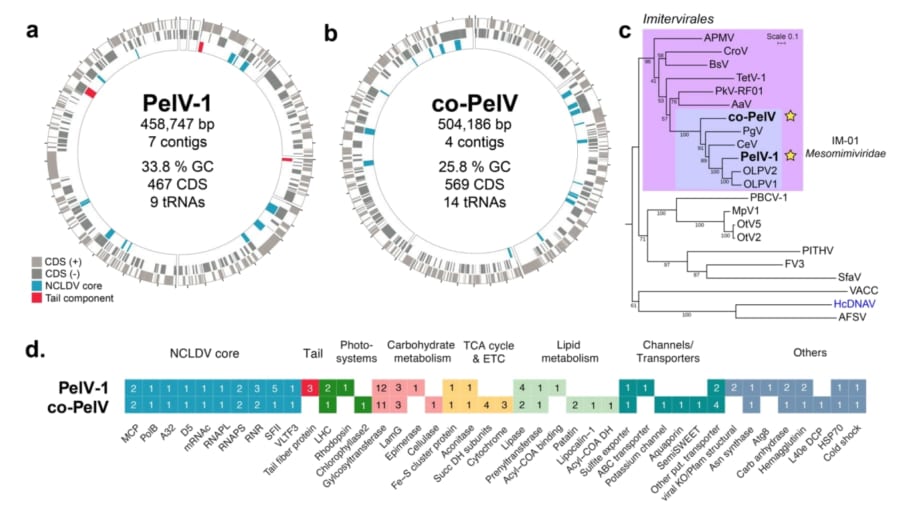
PelV-1のDNAは約46万塩基対という非常に大きなサイズで、これはふつうのウイルスと比べて桁違いに大きいものでした。
その中には、数百にのぼる遺伝子が詰まっていました。
特に注目されたのは、エネルギーを作り出すためのしくみに関係する遺伝子です。
たとえば、細胞の中で食べ物からエネルギーを取り出す「クエン酸回路(TCA回路)」にかかわる酵素の設計図や、脂肪を分解・合成するための遺伝子が見つかりました。
さらに、光を集めるためのたんぱく質(ライトハーベスティングコンプレックス)や、光を感じて反応する「ロドプシン」というたんぱく質の遺伝子もありました。
そのほかにも、水を通す「アクアポリン」、イオンの出入りに関わるチャネル、糖の出入りを助ける輸送体など、さまざまな「便利な道具」がそろっていたのです。
これだけの装備を持つウイルスは、ほとんど例がありません。
研究者たちは、PelV-1がこうした遺伝子をもともとは感染相手のプランクトンから取り入れた可能性があると考えています。
こうした遺伝子が感染した宿主の細胞の動きを変えることで、ウイルスの増殖を助けているのかもしれません。
さらに、PelV-1は感染のときに姿を変える「変身ウイルス」でもありました。
研究チームは、PelV-1が植物プランクトンの細胞に感染するようすを時間を追って観察しました。
すると、ウイルスはまず長いしっぽを使って細胞の表面にピタッとくっついていました。
しかしその後、細胞にしっぽを刺すのではなく、まるごと細胞に取り込まれていったのです。
この動きは「エンドサイトーシス」という、細胞が外のものを飲み込む仕組みに似ているものでした。
もっとおどろいたのは、感染後しばらくたった細胞の中からは、しっぽが付いたウイルスの姿がまったく見えなくなっていたことです。
そのかわり、しっぽのないPelV-1が大量に作られていました。
つまり、ウイルスは感染するときにしっぽを「ポロッと捨ててしまう」ようなのです。
そして、感染し終わったウイルスがまた海の中に放たれると、今度はしっぽがちゃんと生えた姿になっているのです。
まるで「使い捨て変身グッズ」のように、感染のたびにしっぽを付けたり外したりしているのです。
この変身のしくみは、これまでのウイルスではほとんど見られない非常にユニークなものです。
最後に、研究チームはPelV-1とは別に、同じ海水サンプルからもう一種類の巨大ウイルスも見つけました。
こちらは「co-PelV(コ・ペルブイ)」と呼ばれていて、PelV-1と同じグループのウイルスでした。
ただし、このウイルスはPelV-1のような長いしっぽは持っていないようでした。
ではPelV-1のしっぽは何のために存在するのでしょうか?





























![シルバーバック 【釣って遊ぼう!サメまみれ!】 知育玩具 幼稚園 小学校 入園 入学 お祝い プレゼント 準備 ([バラエティ])](https://m.media-amazon.com/images/I/41ejNUfrJZL._SL500_.jpg)